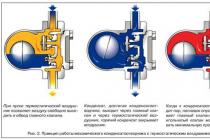
9.5.1. One of the main functions of membranes is participation in the transport of substances. This process is provided by three main mechanisms: simple diffusion, facilitated diffusion and active transport (Figure 9.10). Remember the most important features of these mechanisms and examples of the transported substances in each case.
Figure 9.10. Mechanisms of transport of molecules across the membrane
simple diffusion- transfer of substances through the membrane without the participation of special mechanisms. Transport occurs along a concentration gradient without energy consumption. Small biomolecules - H2O, CO2, O2, urea, hydrophobic low molecular weight substances are transported by simple diffusion. The rate of simple diffusion is proportional to the concentration gradient.
Facilitated diffusion- the transfer of substances across the membrane using protein channels or special carrier proteins. It is carried out along the concentration gradient without energy consumption. Monosaccharides, amino acids, nucleotides, glycerol, some ions are transported. Saturation kinetics is characteristic - at a certain (saturating) concentration of the transferred substance, all carrier molecules take part in the transfer and the transport speed reaches a limiting value.
active transport- also requires the participation of special carrier proteins, but the transfer occurs against a concentration gradient and therefore requires energy. With the help of this mechanism, Na+, K+, Ca2+, Mg2+ ions are transported through the cell membrane, and protons through the mitochondrial membrane. The active transport of substances is characterized by saturation kinetics.
9.5.2. An example of a transport system that performs active ion transport is Na+,K+ -adenosine triphosphatase (Na+,K+ -ATPase or Na+,K+ -pump). This protein is located in the thickness of the plasma membrane and is able to catalyze the reaction of ATP hydrolysis. The energy released during the hydrolysis of 1 ATP molecule is used to transfer 3 Na + ions from the cell to the extracellular space and 2 K + ions in the opposite direction (Figure 9.11). As a result of the action of Na + , K + -ATPase, a concentration difference is created between the cytosol of the cell and the extracellular fluid. Since the transport of ions is non-equivalent, a difference in electrical potentials arises. Thus, an electrochemical potential arises, which is the sum of the energy of the difference in electric potentials Δφ and the energy of the difference in the concentrations of substances ΔС on both sides of the membrane.

Figure 9.11. Scheme of Na+, K+ -pump.
9.5.3. Transfer through membranes of particles and macromolecular compounds
Along with the transport of organic substances and ions carried out by carriers, there is a very special mechanism in the cell designed to absorb and remove macromolecular compounds from the cell by changing the shape of the biomembrane. Such a mechanism is called vesicular transport.

Figure 9.12. Types of vesicular transport: 1 - endocytosis; 2 - exocytosis.
During the transfer of macromolecules, sequential formation and fusion of vesicles (vesicles) surrounded by a membrane occur. According to the direction of transport and the nature of the transferred substances, the following types of vesicular transport are distinguished:
Endocytosis(Figure 9.12, 1) - the transfer of substances into the cell. Depending on the size of the resulting vesicles, there are:
a) pinocytosis - absorption of liquid and dissolved macromolecules (proteins, polysaccharides, nucleic acids) using small bubbles (150 nm in diameter);
b) phagocytosis — absorption of large particles, such as microorganisms or cell debris. In this case, large vesicles are formed, called phagosomes with a diameter of more than 250 nm.
Pinocytosis is characteristic of most eukaryotic cells, while large particles are absorbed by specialized cells - leukocytes and macrophages. At the first stage of endocytosis, substances or particles are adsorbed on the membrane surface; this process occurs without energy consumption. At the next stage, the membrane with the adsorbed substance deepens into the cytoplasm; the resulting local invaginations of the plasma membrane are laced from the cell surface, forming vesicles, which then migrate into the cell. This process is connected by a system of microfilaments and is energy dependent. The vesicles and phagosomes that enter the cell can merge with lysosomes. Enzymes contained in lysosomes break down substances contained in vesicles and phagosomes to low molecular weight products (amino acids, monosaccharides, nucleotides), which are transported to the cytosol, where they can be used by the cell.
Exocytosis(Figure 9.12, 2) - the transfer of particles and large compounds from the cell. This process, like endocytosis, proceeds with the absorption of energy. The main types of exocytosis are:
a) secretion - removal from the cell of water-soluble compounds that are used or affect other cells of the body. It can be carried out both by non-specialized cells and cells of the endocrine glands, the mucosa of the gastrointestinal tract, adapted for the secretion of the substances they produce (hormones, neurotransmitters, proenzymes), depending on the specific needs of the body.
Secreted proteins are synthesized on ribosomes associated with the membranes of the rough endoplasmic reticulum. These proteins are then transported to the Golgi apparatus, where they are modified, concentrated, sorted, and then packaged into vesicles, which are cleaved into the cytosol and subsequently fuse with the plasma membrane so that the contents of the vesicles are outside the cell.
Unlike macromolecules, small secreted particles, such as protons, are transported out of the cell using facilitated diffusion and active transport mechanisms.
b) excretion - removal from the cell of substances that cannot be used (for example, the removal of a reticular substance from reticulocytes during erythropoiesis, which is an aggregated remnant of organelles). The mechanism of excretion, apparently, consists in the fact that at first the excreted particles are in the cytoplasmic vesicle, which then merges with the plasma membrane.
As a rule, it is proteins that are responsible for the functional activity of membranes.
Such proteins include a variety of enzymes, transport proteins, receptors, channels, pore-forming proteins (aquaporins), that is, a variety of protein structures that provide the unique functions of each membrane.
Membrane proteins can be divided into three groups according to their biological role:
I - enzyme proteins with catalytic activity,
II - receptor proteins that specifically bind certain substances,
III - structural proteins.
Enzyme proteins
The most common among all membrane proteins. They include both integral (membrane ATPases) and peripheral (acetylcholinesterase, acid and alkaline phosphatases, RNase) proteins.
Enzymes are large molecules, while the sizes of molecules of substances (substrates) that enter into enzymatic reactions are usually thousands of times smaller. The enzyme interacts with the substrate with a small area of its surface - the active site. The specificity of an enzyme is always determined by the extent to which the surface of its active center corresponds to the surface of the substrate. This principle of structural correspondence is also widely used in the work of cell membrane proteins. In addition, it should be taken into account that the conformation of proteins penetrating the membrane depends on the membrane bilayer, so that their enzymatic activity is also controlled by membrane lipids. This control can be implemented due to both the effect on the affinity for substrates or their availability, and the effect on the life span (strength) of protein associates of membrane enzymes formed in the cell membrane.
Enzymes are part of both plasma and intracellular membranes. For example, on the outer membrane of the epithelial cells lining the digestive organs, there are enzymes that break down nutrients even before they get inside the cell (this process, discovered by the Russian physiologist A.M. Ugolev, is called “membrane digestion”). The outer membrane of liver cells contains more than 20 different enzymes.
Membrane enzymes need to be in contact with their surrounding lipids. When they are removed from the lipid environment (for example, when lipids are extracted from the membrane with non-polar solvents), the work of membrane enzymes is disrupted (the kinetics or the nature of the influence of foreign substances change, or it stops altogether). The activity of such membrane enzymes can be partially restored if lipid micelles are added to them.
An analysis of the nature of lipids that activate membrane enzymes demonstrates the absence of strict specificity - the determining factor is the hydrophilic-lipophilic coefficient of the lipid mixture. In some cases, it is possible to activate a delipidated enzyme even with a detergent. However, such a reactivated enzyme loses the ability to perceive regulatory signals from the outside, which controlled its work in the "living" membrane.
The activating effect of lipids on membrane enzymes can be at least twofold. First, in the presence of lipids, the shape of the membrane enzyme molecule can change, so that its active site becomes accessible to the substrate. Secondly, lipids can play the role of an organizer of an ensemble or conveyor consisting of many enzymes.
Membrane enzyme molecules contain large nonpolar hydrophobic regions. Therefore, in the aquatic environment, they aggregate, due to which most of the active centers are masked. In the presence of lipids, membrane enzymes are organized into ensembles surrounded by annular lipid molecules, and their enzymatic activity can be fully manifested. For the normal operation of membrane enzymes, it is essential that the lipids surrounding them be in a liquid state of aggregation.
Receptor proteins
Receptor proteins are proteins that specifically bind certain low molecular weight substances. When binding specific ligands, receptor proteins reversibly change their shape. These changes trigger chemical responses within the cell. In this way, the cell perceives various signals coming from the external environment and responds to them.
Receptor proteins and proteins that determine the cell's immune response, antigens, can also be both integral and peripheral components of the membrane.
Often, receptors are part of more complex membrane complexes containing executor proteins. For example, the cholinergic receptor receives a signal from a neurotransmitter and transmits it to a channel-forming protein. This reaction opens the membrane permeability to sodium and potassium ions and forms an excitatory potential.
Membrane phospholipids act as a solvent for membrane proteins, creating a microenvironment in which the latter can function. Of the 20 amino acids that make up proteins, six are highly hydrophobic due to the side groups attached to the a-carbon atom, several amino acids are slightly hydrophobic, and the rest are hydrophilic. As we saw in Chap. 5, the hydrophobicity of the peptide groups themselves is minimized upon formation of the a-helix. Thus, proteins can form an integral whole with the membrane. This requires that their hydrophilic regions protrude from the membrane into the cell and out, while the hydrophobic regions penetrate the hydrophobic core of the bilayer. Indeed, those regions of protein molecules that are immersed in the membrane contain a large amount of hydrophobic amino acids and are characterized by a high content of α-helices or α-layers.
Table 42.2. Enzymatic markers of various membranes

The number of different proteins in the membrane varies from 6-8 in the sarcoplasmic reticulum to more than 100 in the plasma membrane. These are enzymes, transport proteins, structural proteins, antigens (i.e., proteins that determine histocompatibility), and receptors for various molecules. Since each membrane is characterized by its own set of proteins, it is impossible to speak of the existence of a certain typical membrane structure. In table. 42.2 shows the enzymatic activities inherent in some types of membranes.
Membranes are dynamic structures. Membrane proteins and lipids are constantly updated. The rates of renewal of different lipids, as well as different proteins, vary over a wide range. The membranes themselves can be renewed even faster than any of their components. This issue will be considered in more detail in the section on endocytosis.
Membrane asymmetry
Asymmetry is an important property of membranes and seems to be partly related to the uneven distribution of proteins in the membrane. Transmembrane asymmetry can also be due to different localization of carbohydrates associated with membrane proteins. In addition, some specific enzymes can be located on the outer or inner side of the membrane; this applies to both mitochondrial and plasma membranes.
The membranes also exhibit local asymmetry. In some cases (for example, in the brush border of mucosal cells), it appears almost at a macroscopic level. In other cases (for example, in the area of gap junctions, tight junctions, and synapses, which occupy a very small part of the membrane area), the areas of local asymmetry are small.
There is also an asymmetry in the distribution of phospholipids between the outer and inner sides of the membranes (transverse asymmetry). So, choline-containing phospholipids (phosphatidylcholine and sphingomyelin) are located mainly in the outer molecular layer, and aminophospholipids
(phosphatidylserine and phosphatidylethanolamine) - mainly in the internal. Cholesterol is usually found in the outer layer in greater quantities than in the inner. Obviously, if such an asymmetry exists in principle, then the transverse mobility (flip-flop) of membrane phospholipids should be limited. Indeed, phospholipids in synthetic bilayers are characterized by an exceptionally low hopping rate - the lifetime of asymmetry can be measured in days or weeks. However, artificial incorporation of certain membrane proteins into synthetic bilayers, for example, the erythrocyte glycophorin protein, the frequency of flip-flop transitions of phospholipids can increase a hundredfold.
The mechanisms of asymmetric lipid distribution have not yet been established. The enzymes involved in the synthesis of phospholipids are localized on the cytoplasmic side of the membranes of microsomal vesicles. Thus, it can be assumed that there are translocases that carry certain phospholipids from the inner layer to the outer one. In addition, specific proteins can be present in both layers, predominantly binding certain phospholipids and leading to their asymmetric distribution.
Integral and peripheral membrane proteins
Most membrane proteins are integral components of membranes (they interact with phospholipids); almost all sufficiently fully studied proteins have a length exceeding 5–10 nm, a value equal to the thickness of the bilayer. These integral proteins are usually globular amphiphilic structures. Both their ends are hydrophilic, and the section crossing the core of the bilayer is hydrophobic. After establishing the structure of integral membrane proteins, it became clear that some of them (for example, molecules of carrier proteins) can cross the bilayer many times, as shown in Fig. 42.7.
Integral proteins are distributed asymmetrically in the bilayer (Fig. 42.8). If a membrane containing asymmetrically distributed integral proteins is dissolved in a detergent and then the detergent is slowly removed, self-organization of phospholipids and integral proteins will occur and a membrane structure will form, but the proteins in it will no longer be specifically oriented. Thus, the asymmetric orientation in the membrane of at least some proteins can be set when they are included in the lipid bilayer. The outer hydrophilic portion of the amphiphilic protein, which is, of course, synthesized inside the cell, must then cross the hydrophobic layer of the membrane and end up outside.

Rice. 42.7. A putative model of the human glucose transporter. It is assumed that the transporter crosses the membrane 12 times. Membrane-crossing regions can form amphiphilic a-helices with amide and hydroxyl side groups and appear to bind glucose or form a channel for its transfer. The amino and carboxyl ends of the chain are located on the cytoplasmic surface. (From Mueckler et al.: Sequence and structure of a human glucose transporter. Science, 1985. 229, 941, with kind permission.)
The molecular mechanisms of membrane organization will be discussed later.
Peripheral proteins do not directly interact with phospholipids in the bilayer; instead, they form weak bonds with hydrophilic regions of specific integral proteins. For example, ankyrin, a peripheral protein, is associated with an integral protein of band III of the erythrocyte membrane. Spectrin, which forms the backbone of the erythrocyte membrane, is in turn bound to ankyrin and thus plays an important role in maintaining the biconcave shape of the erythrocyte. Immunoglobulin molecules are integral proteins of the plasma membrane and are released only together with a small fragment of the membrane. Many receptors for various hormones are integral proteins, and the specific polypeptide hormones that bind to these receptors can thus be considered peripheral proteins. Such peripheral proteins as peptide hormones can even determine the distribution in the plane of the bilayer of integral proteins - their receptors (see below).

1. Cell membrane is an ultra-thin film on the surface of a cell or cell organelle, consisting of a bimolecular layer of lipids with embedded proteins and polysaccharides.
Membrane functions:
barrier - provides selective, regulated passive and active transport.
matrix - provides a certain mutual arrangement and orientation of membrane enzymes relative to substrates in order to implement their optimal interaction.
mechanical - ensures the strength and autonomy of the cell and intracellular structures
2 .Model of the organization today is recognized liquid crystal model. It was first proposed by Singer and Nicholson in 1972. According to this model, the basis of any membrane is a double phospholipid layer. Phospholipid molecules are oriented so that their hydrophilic heads go out and form the outer and inner surfaces of the membrane, and their hydrophobic tails are directed towards the middle of the bimolecular layer. Proteins seem to float in the lipid layer. Surface proteins are located on the outer and inner surfaces of the membrane, being held mainly by electrostatic forces. Integral proteins can penetrate the double layer through. Such proteins are the main component responsible for the selective permeability of cells. membranes.
In addition to phospholipids and proteins, biological membranes contain other chemicals. compounds (cholesterol, glycolipids, glycoproteins).
Functions of membrane proteins. Some membrane proteins are involved in the transport of small molecules across the cell membrane, changing its permeability. Membrane transport proteins can be divided into channel proteins and carrier proteins. Channel proteins contain internal water-filled pores that allow ions (via ion channels) or water molecules to move across the membrane. Many ion channels are specialized for the transport of only one ion; so potassium and sodium channels often distinguish between these similar ions and pass only one of them. Carrier proteins bind each carried molecule or ion and can carry out active transport using the energy of ATP. Proteins also play an important role in cell signaling systems, in the immune response and in cells. cycle.
Functions of membrane carbohydrates. The carbohydrate part of glycolipids and glycoproteins of the plasma membrane is always located on the outer surface of the membrane, in contact with the intercellular substance. Polysaccharides, along with proteins, play the role of antigens in the development of cells. immunity. Participate in the transport of glycoproteins, directing them to their destination in the cell or on its surface.
home function of membrane lipids is that they form a bilayer matrix with which proteins interact.
Lateral diffusion - this is the chaotic thermal movement of lipid and protein molecules in the plane of the membrane. With lateral diffusion, adjacent lipid molecules jump around and, as a result of such successive jumps from one place to another, the molecule moves along the membrane surface. The root-mean-square movement of molecules during diffusion during time t can be estimated by the Einstein formula: Skv = 2cor(Dt), where D is the coefficient. lat. diffusion of the molecule.
Lateral diffusion of integral proteins in the membrane is limited, this is due to their large size and interaction with other membranes. proteins. Membrane proteins do not move from one side of the membrane to the other ("flip-flop" jumps), like phospholipids.
The movement of molecules on the surface of the cell membrane during the time t is determined experimentally by the method of fluorescent labels - fluorescent molecular groups. It turned out that the root-mean-square movement of a phospholipid molecule per second along the surface of the erythrocyte membrane corresponds to a distance of 5 μm, which is comparable to the size of cells. A similar value for protein molecules was 0.2 µm per second.
The coefficients calculated according to the Einstein formula lat. diffusion for lipids 6 * 10 (at -12 st) kV.m / s, for proteins - about 10 (at - 14 st) kV.m / s.
The frequency of molecule jumps from one place to another due to lat. diffusion can be found by the formula:
v = 2kor(3D/f),
where f is the area occupied by one molecule on the membrane.
artificial membrane usually it is a rigid selectively permeable partition separating the mass transfer apparatus into two working zones, in which different pressures and compositions of the mixture being separated are maintained. The membranes can be made in the form of flat sheets, pipes, capillaries and hollow fibers. Membranes line up in membrane systems. The most common artificial membranes are polymeric membranes. Under certain conditions, ceramic membranes can advantageously be used. Some membranes operate in a wide range of membrane operations such as microfiltration, ultrafiltration, reverse osmosis, pervaporation, gas separation, dialysis, or chromatography. The method of application depends on the type of functionality incorporated into the membrane, which may be based on dimensional isolation, chemical affinity, or electrostatics.
Liposomes. or phospholipid vesicles (vesicles), usually obtained by swelling dry phospholipids in water or by injecting a solution of lipids into water. In this case, self-assembly of the bimolecular lipid membrane occurs. In this case, all nonpolar hydrophobic tails are inside the membrane and none of them comes into contact with polar water molecules, however, more often spherical multilamellar liposomes are obtained, consisting of several bimolecular layers - multilayer liposomes.
The individual bimolecular layers of the multilayer liposome are separated by an aqueous medium. The thickness of the lipid layers is, depending on the nature of the lipids, 6.5 - 7.5 nm, and the distance between them is 1.5 - 2 nm. The diameter of multilayer liposomes ranges from 60 nm to 400 nm or more. Single-layer liposomes can be obtained by various methods, for example, from a suspension of multi-layer liposomes, if they are treated with ultrasound. The diameter of single-layer liposomes is 25 - 30 nm. Liposomes are in some way the prototype of the cell. They serve as a model for studying various properties of cl. membranes. Liposomes have also found direct application in medicine (phospholipid microcapsule for drug delivery to certain organs and tissues). Liposomes are non-toxic, completely absorbed by the body.
By clicking on the "Download archive" button, you will download the file you need for free.
Before downloading this file, remember those good essays, control, term papers, theses, articles and other documents that are unclaimed on your computer. This is your work, it should participate in the development of society and benefit people. Find these works and send them to the knowledge base.
We and all students, graduate students, young scientists who use the knowledge base in their studies and work will be very grateful to you.
To download an archive with a document, enter a five-digit number in the field below and click the "Download archive" button
Similar Documents
The structure of biological membranes and the structure of their basis - bilipid layer. Molecular weight of membrane proteins, their difference in the strength of binding to the membrane. Dynamic properties of biological membranes and the importance of organization for biological systems.
abstract, added 12/20/2009
The structure of membranes. erythrocyte membranes. myelin membranes. chloroplast membranes. Inner (cytoplasmic) membrane of bacteria. Virus membrane. Membrane functions. transport across membranes. Passive transport. active transport. Ca2+ is a pump.
abstract, added 03/22/2002
Proteins and lipids as the main components of membranes. Phospholipid composition of subcellular membranes of rat liver. Long hydrocarbon chains. Gram-positive bacteria membranes. Pathways for the biosynthesis of membrane lipids and mechanisms for their delivery to their destinations.
abstract, added 07/30/2009
The abundance and complexity of the structure of internal membranes as one of the main features of all eukaryotic cells. The concept, properties and functions of membranes: barrier, transport. The essence and purpose of ion and calcium channels, methods from the study.
abstract, added 10/19/2014
Functions of biological membranes and their components. Spectroscopic methods for measuring the rate of rotation of lipids and proteins inside the membrane and the rate of lateral diffusion of these components in the plane of the membrane. Use of spin or fluorescent probes.
abstract, added 08/01/2009
Diversity and role of membranes in the functioning of prokaryotic and eukaryotic cells. Morphology of membranes, their isolation. X-ray diffraction, electron microscopy. Destruction of cells, separation of membranes. Purity criteria for membrane fractions.
term paper, added 07/30/2009
Ultrastructure of biological and molecular structure of cytoplasmic membranes, their main functions. The physical nature of the forces of interaction between proteins and lipids in their structures. Methods for studying and researching artificial models of cytoplasmic membranes.